Il Sindaco di Ponza denuncia chi approda all’Isola di Palmarola per documentare il crescente bracconaggio e caccia illegale della zona
Aprile 2016
Il Sindaco di Ponza denuncia alla polizia municipale il sottoscritto (reporter di natura) e Andrea Rutigliano(volontario CABS). Eppure quello che abbiamo visto e fotografato parla chiaro: l’isola di Palmarola è rifugio di cacciatori illegali e bracconieri in questo periodo (migrazione di avifauna). In Italia la caccia in primavera è vietata dal 1992 !!!!!
Report (Portfolio) Palmarola 2016 su www.paorossi.it
Andrea mi guarda e mi dice “qui ci sono le piazzole da trappola ma non ci sono trappole, i bracconieri hanno saputo del nostro arrivo e le hanno rimosse in tempo”. Un’ora dopo però girovagando e cercando prove di caccia illegale su Palmarola vedo in lontananza Andrea che raccoglie qualcosa sui bordi di un dirupo a picco sul mare…” eccola! è una trappola! Un gabbiano deve averne rubata una prima che le rimuovessero, l’ha portata qui e si è mangiato l’uccellino che vi era intrappolato”. La trappola è recente, il Gabbiano ha smascherato i bracconieri.
(PARTE3di3)
Le preoccupazioni del CABS erano fondate !!!
L’isola di Palmarola (Ponza) liberata dai bracconieri
(…) sequestrati tre fucili, di cui due di provenienza furtiva e uno con matricola abrasa, 111 trappole, 71 cartucce; un cacciatore arrestato, una altro denunciato, altri sono rimasti nascosti negli anfratti. E’ questo il bilancio dell’operazione condotta dal Nucleo investigativo della Forestale di Latina nella mattinata di oggi. (…)
(…) La denuncia sporta presso la stazione dei carabinieri di Ponza da parte del Cabs ha avuto un seguito, e messo in campo una straordinaria macchina di collaborazione tra l’Arma e la Forestale (…)
Su www.paorossi.it le immagini che hanno contribuito a dimostrare la presenza di caccia illegale e bracconaggio sull’isola.
(Torriglia, 19 Apr 16) Presso la sala consiliare del Comune di Valbrevenna (località Molino Vecchio) si terra un incontro con la popolazione locale, riguardo la presenza costante del Lupo nelle nostre valli. In particolare l’incontro mira a rispondere alle numerosissime domande e preoccupazioni poste dal vasto pubblico riguardo alla ipotetica pericolosità del predatore, alla sua provenienza, alla sua alimentazione ed alle sue abitudini.
Coordineranno: Daniela Segale, Antonio Federici, Adriano Zanni (Parco Antola).
Interverranno diversi esperti del settore (Renato Cottalasso, Roberto Sobrero, Massimo Campora) e operatori foto/video (Gabriele Cristiani, Paolo Rossi).
Dino è un membro del C.A.B.S. che spesso trascorre le ferie sull’isola di Cipro “cercando di farsi ammazzare”: infatti Dino combatte il bracconaggio nella parte greca dell’isola, “dove la tradizione (reti) è diventata quasi estinzione (reti + richiami elettronici)” allo scopo di catturare piccoli uccelli migratori (ma anche gli uccelli rapaci che finiscono in trappola, come Gufi e Falchi vengono uccisi perché non utili). L’estinzione della fauna selvatica a Cipro come in molte parti del mondo è un business: cucina, medicina, tradizione, ecc…. e chi si “intromette” rischia la propria incolumità.
Centinaia di miliaia di uccelli migratori e stanziali vengono ammazzati ogni anno a Cipro parte greca
Dino libera uno Sparviere dopo le cure, era in una rete e sarebbe stato ucciso dal bracconiere cipriota
Dino e altri membri del CABS rischiano la propria vita per impedire la possibile estinzione di alcune specie di uccelli
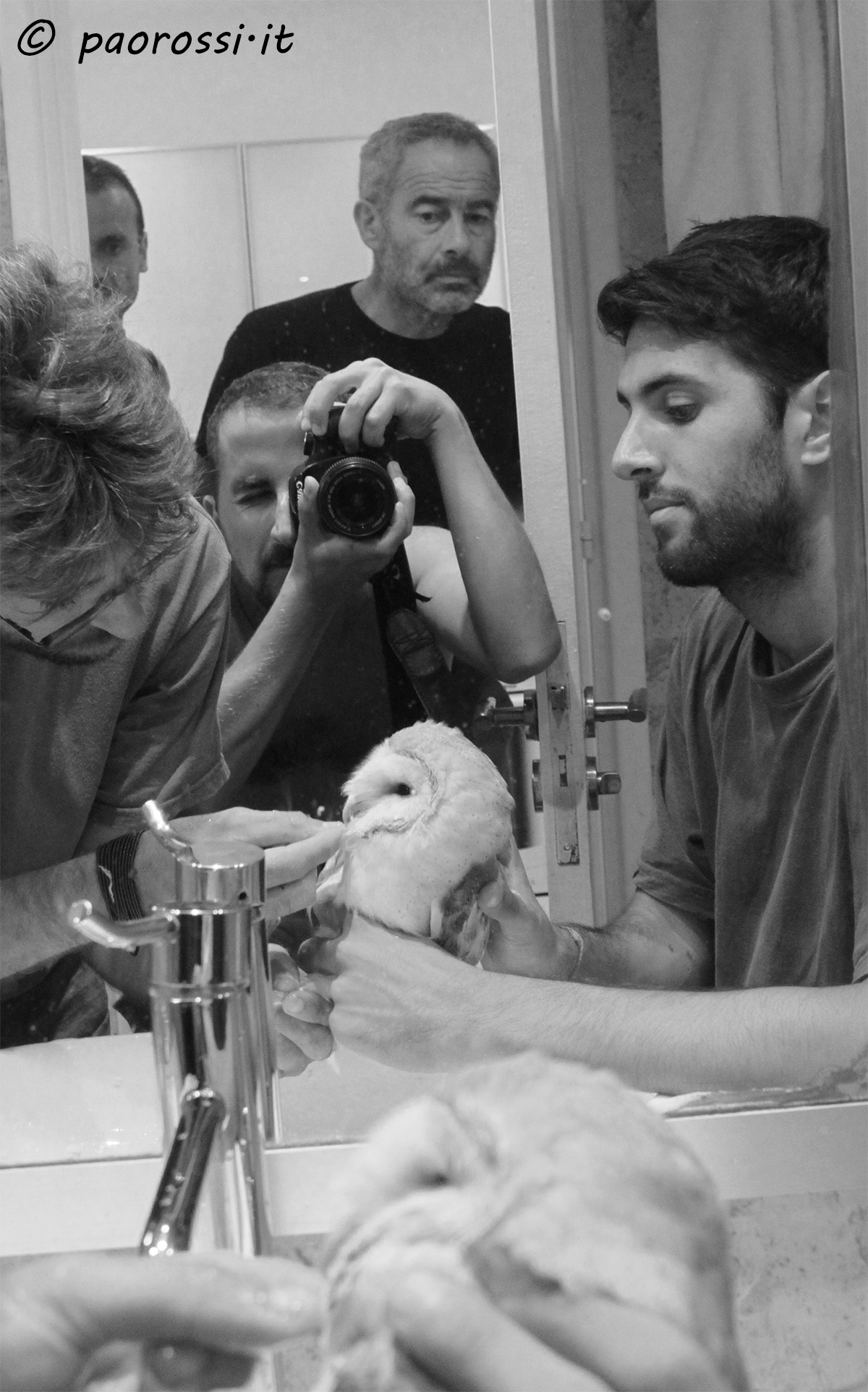
Le trappole non fanno selezione: in foto un Barbagianni pulito dal vischio di una trappola cipriota. Tornerà in libertà qualche giorno dopo lo scatto
Le reti unite ai richiami elettronici rappresentano un mix terrificante (tradizione+tecnologia moderna) per gli uccelli migratori spossati dal lungo viaggio
Come preservare la Biodiversità attraverso le nostre scelte di vita quotidiana.
Cowspiracy
L’impatto devastante e “determinante” degli allevamenti zootecnici sul pianeta e i motivi per i quali le grandi associazioni ambientaliste non parlano/combattono il problema
Il Sale della Terra
Storia e Attivismo di un grandissimo fotografo del nostro tempo
Sorpresa: ho ripescato stralci di vecchiotti ma sempre attuali articoli dell’Internazionale
Tratto da Internazionale 23 maggio 2008
Qui pianeta Terra di Michael Pollan
Il virus del cambiamento (…) I motivi per non fare nulla sono molti e apparentemente convincenti, almeno per chi ha la mentalità dell’energia a basso costo. Vorrei suggerire alcuni motivi da mettere sull’altro piatto della bilancia. Se facciamo qualcosa, diamo l’esempio ad altri. Se un numero sufficiente di persone cominciasse a fare qualcosa, ognuna influenzerebbe altre persone e si verificherebbe una reazione a catena di comportamenti diversi. Il mercato dei prodotti biologici e delle tecnologie alternative crescerebbe. Molti prenderebbero coscienza del problema, forse cambierebbero. Nascerebbero nuovi imperativi morali e nuovi tabù: forse guidare un Suv, mangiare una bistecca da un chilo o illuminare una casa come fosse un aeroporto sarebbero considerati comportamenti incoscienti. Non possedere troppe cose potrebbe diventare di moda. E i pionieri del nuovo stile di vita avrebbero il diritto morale di chiedere anche agli altri – persone, industrie, paesi – di cambiare i loro comportamenti. Tutto questo, in teoria, potrebbe succedere. Si tratta di un cambiamento sociale di tipo virale, cioè non lineare e non prevedibile. Chissà, forse il virus arriverà fino a Chongging e contagerà anche “il mio gemello cattivo cinese”. O forse succederà il contrario, e tra qualche anno l’ambientalismo sarà passato di moda. Scegliere di vivere in modo più ecologico è solo una scommessa, niente di più e niente di meno. Ma forse, anche se le probabilità di vincerla sono poche, dovremmo accettarla tutti. A volte bisogna agire come se le nostre azioni potessero fare una qualche differenza, anche se non ne siamo certi. Dopotutto, è proprio questo che è successo nella Cecoslovacchi e nella Polonia comuniste, quando gli uomini come Vaclav Havel e Adam Michnik hanno deciso di vivere “come se” la loro fosse un società libera. Quella scommessa improbabile creò un piccolo spazio di libertà, che con il tempo si allargò fino a provocare il crollo di tutto il blocco orientale. Secondo Vaclav Havel, di fronte alla crisi ambientale la gente dovrebbe cominciare a comportarsi “come se dovesse vivere su questa Terra per sempre e un giorno dovesse rendere conto delle condizioni in cui la lascia”. Sono d’accordo, ma vorrei dare un suggerimento meno astratto e scoraggiante: trovate una cosa da fare nella vostra vita che non sia spendere né votare, che forse sconvolgerà il mondo come un virus o forse no, ma che sia reale e specifica (oltre che simbolica) e che, qualunque cosa succeda, abbia i suoi vantaggi. Potreste decidere di rinunciare alla carne, cosa che ridurrebbe di un quarto la vostra impronta di carbonio. Oppure per un giorno alla settimana, astenetevi completamente da qualsiasi attività economica: niente shopping, niente automobile, niente elettronica.
Tratto da Internazionale del 28 novembre 2008
Sulla via del disastro di Herman Daly
(…) Limite oltrepassato – Le dimensioni dell’economia globale si stanno avvicinando al limite massimo di sopportazione del pianeta. Man mano che gli oceani si svuotano di pesci, le foreste si riducono e i gas serra nell’atmosfera aumentano, i costi ambientali e sociali di un’ulteriore crescita sono destinati a intensificarsi fino al punto in cui il prezzo che pagheremo per ogni punto extra di crescita sarà maggiore dei benefici che ne ricaviamo. Anzi, ci sono elementi per sostenere che abbiamo già oltrepassato quel limite, almeno in paesi ricchi come gli Stati Uniti e la Gran Bretagna. Fino quando il nostro sistema economico si baserà sulla rincorsa alla crescita, andremo incontro a un disastro ambientale ed economico. Per evitarlo, dobbiamo spostare l’obiettivo dalla crescita quantitativa allo sviluppo qualitativo e porre limiti rigorosi alla velocità con cui consumiamo le risorse della Terra. Il valore dei beni prodotti può comunque aumentare, ma le dimensioni fisiche dell’economia devono essere contenute entro un livello che il pianeta sia in grado di sostenere. Dopo duecento anni di crescita è difficile immaginare come possa essere un’economia controllata, ma non vuol dire necessariamente che ci congeleremo al buio sotto il giogo di una tirannia comunista. Gran parte dei cambiamenti possono essere introdotti gradualmente.
Meglio vivere senza stress di Kate Soper (…) sempre più persone di rendono conto che forse la vita non consiste solo nel lavorare per spendere, e provano a semplificarla e a rivedere i loro valori e desideri. Se passassimo in massa a un’economia in cui si lavora meno, il numero di persone, merci e informazioni costrette a circolare sarebbe minore e questo ridurrebbe anche il consumo di risorse e le emissioni di carbonio. Le persone avrebbero più tempo da dedicare a se stesse e alla famiglia. E in una crisi economica come quella attuale i politici potrebbero ottenere più collaborazione e rispetto da parte dell’elettorato se si impegnassero su questi temi.
Giunto nel grande Altipiano abruzzese noto dei corvi imperiali, sembrano in corteggiamento. A volte seguono i lupi per approfittare dei loro “avanzi” di cibo (Corvus Corax).
Resti di predazione, feci e le loro impronte fresche sul fango “di una pista di cervo”: i lupi sono presenti in questa zona e forse, mi stanno osservando la nuca.
Non mi resta che prepararmi un giaciglio comodo e affacciato su una grande radura, sperare che il vento sia con me e attendere i principi delle tenebre (Canis lupus italicus).
E dopo giorni e ore di attesa … eccolo, è un maschio adulto parzialmente affetto da rogna (forse favorita da quest’inverno caldo-umido): è quasi buio e il suo trotto è molto rapido: sembra una danza selvaggia e forse per lui, la caccia è appena iniziata!
L’Abruzzo è una delle zone italiane con il patrimonio di biodiversità più ricco. Infatti in questa regione i lupi (e non solo!) sono numerosi. Il Parco Nazionale di Lazio Abruzzo e Molise tra gli altri, è famoso per le sue straordinarie faggete.
Il drammatico inverno anomalo causato dal surriscaldamento globale, permette perlomeno di vedere integralmente questi straordinari faggi vetusti (Fagus sylvatica).
La cementificazione nelle aree verdi di tutta Italia è impressionante. Anche nei Comuni delle zone di Parco ciò avviene (foto sotto 2016 Opi – Parco Nazionale Lazio Abruzzo e Molise), anche se in modo più circoscritto e subdolo.